 |
 |
 |
|
|
|
|
|
| 日本語 | 検索表へ | English | Go to Key Tab. | ||
日本産花粉の標徴Ⅰ・Ⅱ 改訂
中村 純・守田 益宗・黒田 登美雄
 |
 |
Revision of Nakamura's(1980) diagnostic characters of pollen grains of Japan
By Jun Nakamura, Yoshimune Morita and Tomio Kuroda
Preface
It goes without saying that it is important to understand the morphology of pollen in any field where pollen is the subject of research. Sketches and photographs are useful for pollen grain identification and, in Japan, several pollen pictorial guide books have been published, the first being the sketch collections of Jimbo (1933) and Yamazaki (1933). A later work, Diagnostic characters of pollen grains of Japan Part I and II, published in 1980 by the late Dr. Jun Nakamura, our teacher, contains optical micrographs and descriptions of approximately 1,100 types of pollen. Many researchers, especially pollen analysts, have benefited from the use of this publication. Professor Nakamura hoped that this pollen pictorial guide book would be widely available, not only to domestic and foreign researchers but also to students aspiring to study pollen; however, copies of this publication are already difficult to obtain. Therefore, before Dr. Nakamura's research materials are lost, we decided to publish this pictorial pollen guide to attract the will of our teacher.
This revised and updated version of Professor Nakamura’s publication has been compiled based on the following objectives:
1) The addition of a new pollen key table because it may be difficult to distinguish pollen with similar shapes only by comparing the plates.
2) Use of the APG III classification system that emphasizes the molecular lineage of plants in place of the traditional Engler system that emphasizes plant morphology.
3) The development of an online version of the pictorial pollen guide that makes it possible to search and browse the content on the Web.
4) The addition of English text.
In addition, based on new findings, we have revised and corrected erroneous descriptions contained in the 1980 publication.
This pictorial pollen guide was co-edited by Morita and Kuroda. Morita created the pollen key table and revised the descriptions, and Kuroda translated the text into English and created the website.
Pollen treatment method and the plate and pollen description terminology
The pollen samples were treated using the acetolysis method of Erdtman (1943) and were sealed using non-volatile silicon oil (6,000 cs). The sealing method (applied on the glass slides) was based on Andersen (1960).
Micrographs were taken using a Nikon Apophoto type microscope (objective lens PlanApo 100 × 160 mm/NA1.35, projection lens, eyepiece CF photo 8 ×) with an HFM-type imaging device. The film was developed using Kodak High Contrast Copy film (5069) with the DK-20 formulation. This film was read using a film scanner at a resolution of 2,400 dpi and saved as JPEG image data.
As a general rule, the scale of the plates is 10 μm; scales that differ from this are shown individually.
Pollen grain measurements
To determine pollen grain size, 50 grains of each species were measured, and the maximum and minimum values recorded, as documented in the descriptions. Where the polar length and the equatorial diameter differ, the values are presented as the equatorial diameter x the polar length. For pollen with unclear poles, the maximum diameter is presented; the minimum diameter is also provided, if necessary. An eyepiece micrometer and a digital micrometer (Kogaku Co., Ltd.) were used to measure the pollen grain size.
The density per unit area of protrusions and meshes is provided in the descriptions, where relevant. As a general rule, the density values were determined on the central part of the proximal pole surface for monocolpate pollen, and as the number of protrusions per 5 × 5 (25 μm2) or 10 × 10 (100 μm2) areas on the mesocolpium for other types of pollen. In principle, where the thickness of the exine was not uniform, it was measured on the proximal pole surface or on the mesocolpium.
Plant species considered
The pollen types described are nearly limited to those that could be pollen-analyzed, and pollen types with an extremely low likelihood of becoming fossil pollen, such as pollen of the Orchidaceae, and Metaplexis, have been excluded.
The scientific names and arrangement of the plant species, the pollen of which is described and illustrated in the micrographs, is based on Ohi (1978) in Nakamura (1980). This arrangement follows the format of the APG III classification system by Yonekura and Murata (2013), and the scientific names follow Yonekura and Kajita (2003-).
Exine structure
Pollen grains are mostly covered by an intine consisting of pectin and cellulose and an exine consisting of sporopollenin. However, when the acetolysis treatment is performed, only the chemically tough exine remains. The exine is divided into the endexine and ektexine*, and the ektexine is further divided into three layers: foot, columellae, and tectum (Fig. 1). A layered exine structure is not found in gymnosperm pollen, but is common in the pollen produced by many angiosperms. Pollen grain shape is determined as a result of various changes in the exine and the germination part (the germinal aperture, which is the outlet of the pollen tube), in relation to the pollination mode and evolution.
* Synonymous with ectexine (Punt et al., 2007)
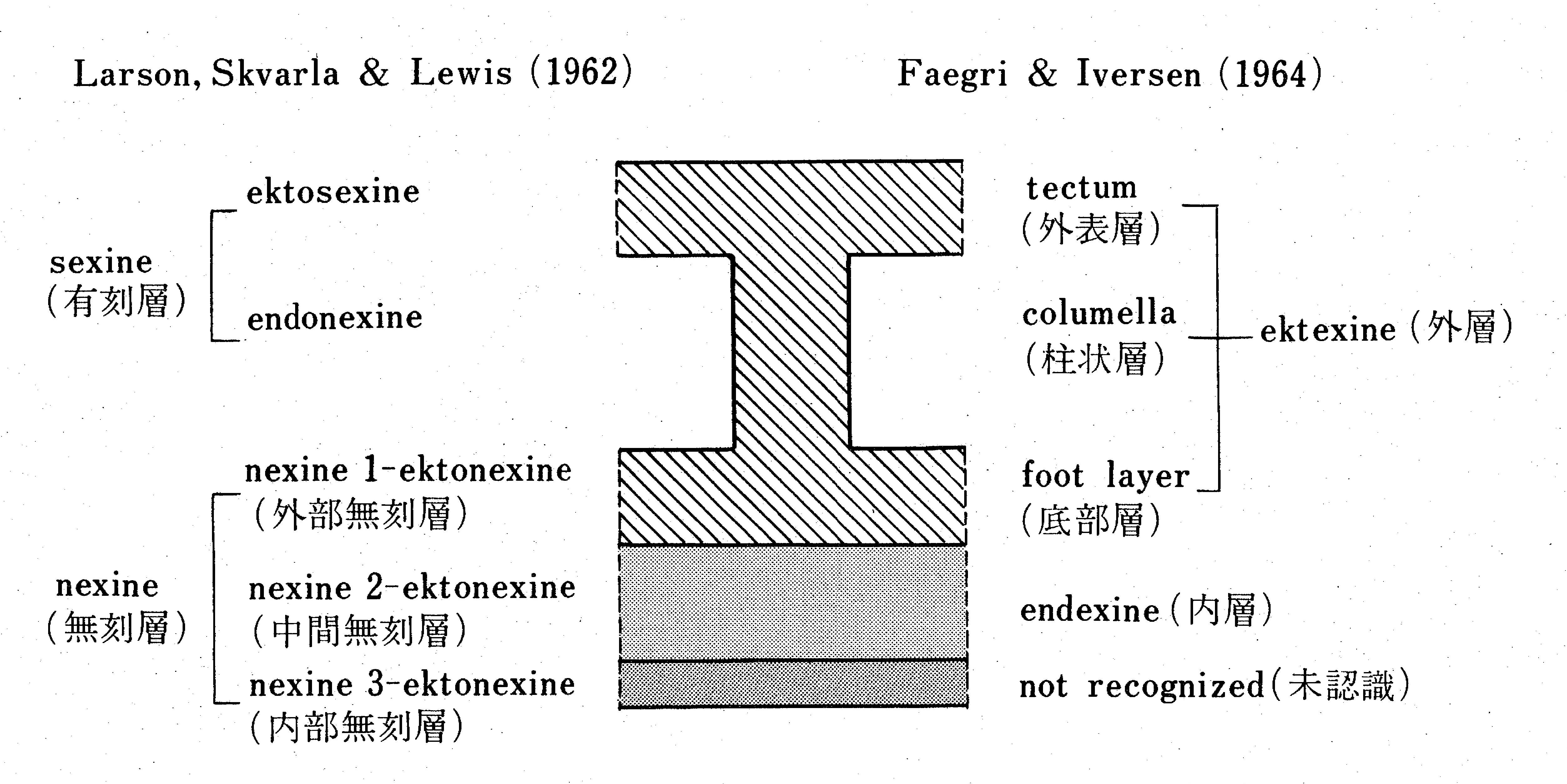
Fig.1: A diagrammatic representation of a stratification of the pollen grain exine.
Pollen morphology
The morphological characteristics of pollen grains vary among plant types; however, the following four morphological features are key characteristics when identifying pollen: 1) pollen grain size; 2) shape; 3) number, position, and arrangement, and shape and structure of the germinal apertures; and 4) pollen sculpture and the associated structures.
Pollen morphology, particularly grain shape and position of structures, cannot be accurately described unless related to a direction of observation, and pollen grains are usually described in terms of polar and equatorial orientations, similar to those applied to the earth. Pollen mother cells divide to form four pollen cells that develop into pollen grains. During this development, each cell is arranged three-dimensionally or on a plane. When the four pollen grains are together, the central direction is termed the proximal axis, and the opposite direction is termed the distal axis along the axis (polar axis) toward the center. The axis that vertically intersects the central part of the polar axis is the equatorial axis.
1) Size
Pollen grain size is an important characteristic for distinguishing between pollen types, for example, where the outer shape of pollen grains is the same but the size is substantially different. Therefore, it is necessary to indicate the grain size in morphological descriptions of pollen. In some cases, the size of the germinal aperture and the size of the protrusions of the sculpture are also important distinguishing features. During the preparation of pollen for analysis, the size of the grains changes depending on the treatment method used, the sealing medium, and the time after sealing. In general, treated pollen grains tend to be larger when the sealing medium used does not require dehydration compared to the size when a sealing medium is used that does require dehydration. Use of the acetolysis method or the KOH method when preparing pollen for analysis also influences the grain size. Reitsma (1969) and Meltsov et al. (2008) report the differences in pollen grain size due to differences in the treatment methods and sealing media used. Because silicon oil was used as the sealing medium when preparing pollen for analysis for this pictorial pollen identification guide, the possibility of a change in grain size over time can be ignored.
2) Shape
In some plants, when pollen grains are formed, the individual grains do not separate and remain bound in tetrads. Such pollen is termed multi-aggregated pollen, and this state of aggregation is an important distinguishing feature in pollen identification.
Some gymnosperms produce pollen grains with structures termed air sacs. The pollen of each of these gymnosperm species has particular characteristics relating to the size and shape of the airs sacs and how they adhere to the corpus.
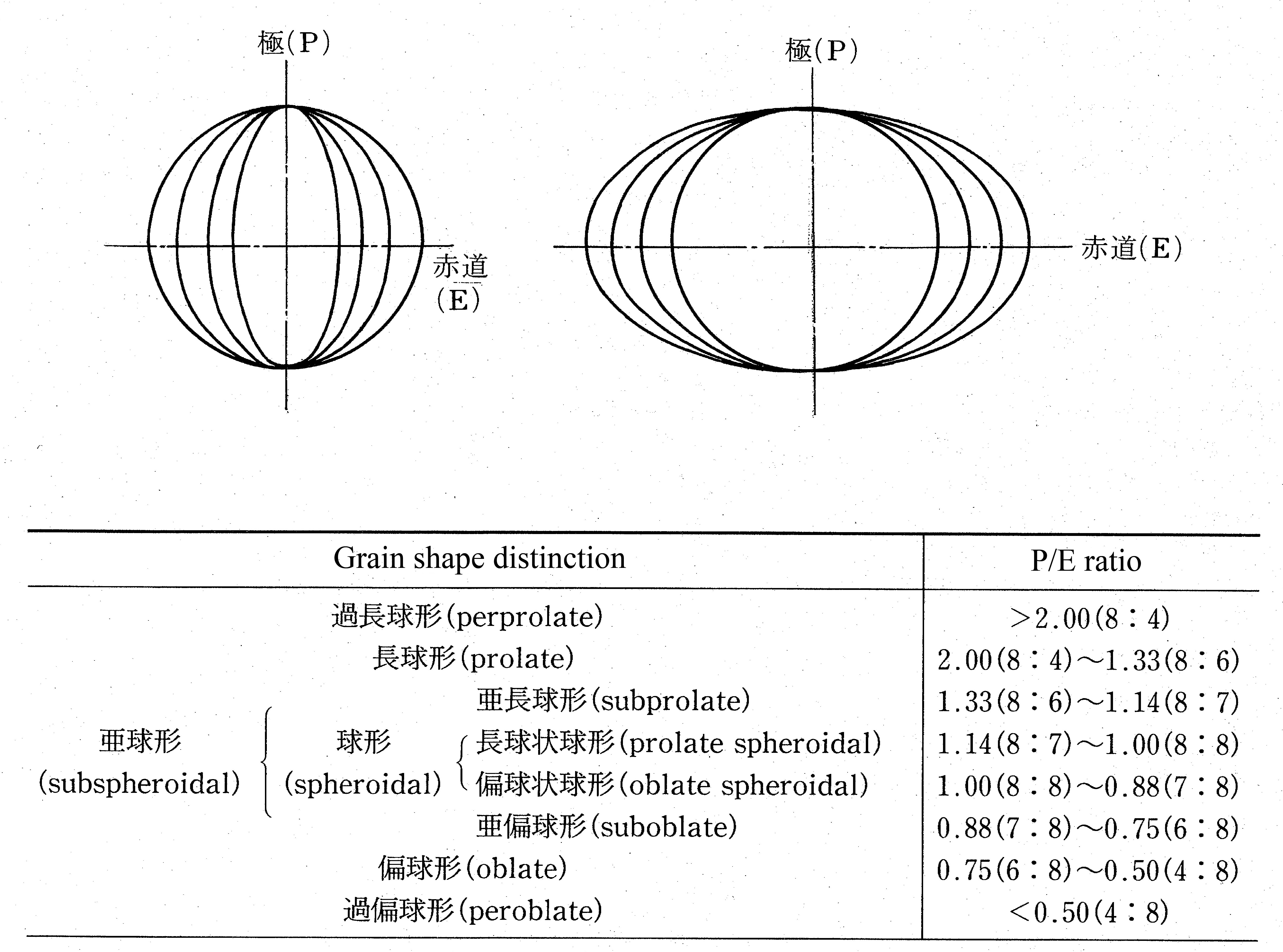
Fig. 2 : Pollen grains in shape classes (P/E ratio) according to Erdtman's(1952)
classification.
Most pollen occurs as single grains, and different shapes can be observed for a single grain, depending on the viewing direction. When viewed from the polar direction, termed the polar view, various shapes can be recognized, depending on the shape of the contour. When viewed from the equatorial direction, termed the equatorial view, various shapes from perprolate to peroblate can be distinguished by the ratio of the length in the polar direction (P) to that in the equatorial direction (E), that is, the P/E ratio (Fig. 2). Because the polar view and the equatorial view change with the sealing method used, and aging, and because it is not always possible to observe pollen grains from the ideal direction, it is often necessary to make corrections, based on experience, when actually observing and recording pollen grain shape.
3) Apertures
The germinal aperture is comprised of a pore and a furrow, according to its shape. The germinal aperture refers to a circular or elliptical shape with a minor to major axis ratio of 2 or less. The furrow refers to a crevice shape, sharp at both ends, with a minor to major axis ratio of 2 or more. The aperture is divided into the ectoaperture in the ectexine and the endoaperture in the endexine, and is visually distinguished into three types: porate, colpate, and colporate, according to the shape and combination of the ectoaperture and the endoaperture. In addition, the aperture is usually further divided into types based on the structure of the ectexine and endexine around the pore. The number, position and arrangement, and shape and structure of the germinal aperture are important distinguishing features because they are generally fixed within each taxon. Most pollen grains that have multiple germinal apertures have these arranged on the equatorial plane or symmetrically on the equatorial plane; however, in some taxa, such as Cyperaceae, the pollen grains exhibit heteropolarity. In gymnosperms, the exine on the distal pole surface is often thin, and is the place where pollen tubes emerge during germination. This area of thin exine is termed the leptoma.
4) Sculpturing
Pollen grains show a range of types of sculpturing, identified based on the structure and components of the exine, as viewed in cross section. Regarding the ectexine, there are types of pollen grains that have a tectum above the columellae (termed tectate) and types of grains that lack a tectum (termed intectate). In tectate pollen grains, different sculpturing is often observed when focusing on the top of the tectum compared to when focusing on the columellae. Therefore, when observing pollen grain sculpture under a microscope, it is important to carefully observe the exine cross section and perform LO analysis by adjusting the focus (Erdtman, 1952).
About the key and the terms used
Pollen grains have various shapes and sculptures and the terms that describe pollen morphology have been defined by many palynologists. The classification of pollen types used in this pictorial pollen guide is based on 24 types, following Faegri and Iversen (1975). The morphological terminology is drawn from the Dictionary of palynology (Palynological society of Japan, 1994), the Glossary of pollen and spore terminology (Punt et al., 2007), and the following documents.
We would like to express our cordial thanks to Professor Emeritus Norio Miyoshi of Okayama University of Science, Professor Kee-Ryong Choi of Ulsan University of Korea, and Curators of the Museum of Natural History of Osaka for their assistance in the publication of this revised and updated version of Professor Nakamura’s pictorial pollen guide.
References
Andersen, S. Th.(1960)Silicone Oil as a Mounting Medium for Pollen Grains. Danmarks Geologiske Undersøgelse IV. Række, vol. 4, 1–24.
Erdtman, G(1943)An introduction to Pollen Analysis. Chronica botanica Company, Waltham.
Erdtman, G. (1952) : Pollen Morphology and Plant Taxonomy. Almqvist & Wiksell, Stockholm.
Faegri, K.and J. Iversen (1975) : Textbook of modern pollen analysis. 3rd Ed. Munksgaard, Copenhagen.
Jimbo, T.(1933): The Diagnoses of the pollen of Forest Trees. 1. Sci. Rep.Tohoku Univ 4th. ser. (Biol), vol.8 287-299.
Larson, D. A., Skvarla, J. J., and Lewis, C. W., (1962): An electron microscope study of exine stratification and fine structure. Pollen et Spores, vol. 4, 233–246.
Meltsov, V., Poska, A. & Saar, M.(2008): Pollen size in Carex: The effect of different chemical treatments and mounting media. Grana, vol. 47, 220-233.
Nakamura, J. (1980) Diagnostic Characters of Pollen Grains of Japan. Part 1&2. Special Publications from the Osaka Museum of Natural History, vol. 12&13. Osaka Museum of Natural History, Osaka (in Japanese).
Palynological society of Japan (1994) : Dictionary of palynology. Asakura-Shoten, Tokyo (in Japanese).
Ohwi, J. (1978) : Flora of Japan. a revised and enlarged edition. Shibundo, Tokyo (in Japanese).
Punt, W., Hoen, P.P., Blackmore, S., Nilsson, S. and Thomas, A. Le (2007) : Glossary of pollen and spore terminology. Review of Palaeobotany and Palynology, vol. 143, 1–81.
Reitsma, T. (1969) : Size modification of recent pollen grains under different treatments. Review of Palaeobotany and Palynology, 9, 175–202
Yamazaki, T. (1933): Pollen and spore morphology in Sakhalin research forest, Kyoto Imperial University. Bulletin of the Kyoto Imperial University Forests, No.5, 1-27.
Yonekura, K. and Kajita, T. (2003-) : BG Plants Japanese-Scientific name index (YList). http://ylist.info
Yonekura, K. and Murata, J. (2013) : Syllabus of the vascular plants of Japan. Hokuryukan, Tokyo (in Japanese).
--------------------------------------------------
日本産花粉の標徴Ⅰ・Ⅱ 改訂
中村 純・守田 益宗・黒田 登美雄
はじめに
花粉を研究対象とするどの分野であっても,まずは,花粉の形態の理解が重要なのは言うまでもない.花粉粒の鑑定にはスケッチや写真などが便利である.そのため,我国でもJimbo(1933),山崎(1933)のスケッチ集にはじまり,いくつかの花粉図鑑が世にだされてきた.なかでも,私たちの恩師である故・中村純先生が1980年に出版された「日本産花粉の標徴Ⅰ・Ⅱ」は約1,100種に及ぶ花粉の光学顕微鏡写真と記載文からなり,多くの研究者,特に花粉分析研究者がその恩恵をうけてきた.中村先生は,同書が国内・外の研究者だけでなく,花粉研究を志す学生にも広く利用できることを願っていたが,同書はすでに入手困難である.そこで,中村先生の研究資料が散逸する前に,その意思を引く次ぐべく,本花粉図鑑を出版することとした.
本花粉図鑑は以下の方針のもとにまとめた.
(1)形がよく似た花粉ではそれらの図版の比較だけでは区別が難しいこともあることから,花粉の検索表を新たに付す.
(2)旧来の植物形態を重視したエングレル(Engler)体系ではなく分子系統を重視したAPG分類体系に変更する.
(3)WEB上で検索・閲覧できるようにする.
(4)英文を付す.
なお,旧書中の誤記載や新知見に基づく訂正もあわせて行った.そして,本花粉図鑑は守田と黒田の共同編集であるが,検索表と記載文は守田が,英文およびWEB作成は黒田がそれぞれ分担した.
花粉の処理法と図版および花粉記載
使用した花粉はErdtmanのacetolysis法(1943)により処理し,不揮発性のsilicon oil(6,000 cs)で封入した.封入法はAndersen(1960)に準拠した.
顕微鏡写真はNikon Apophoto型顕微鏡(対物レンズ PlanApo 100x 160/1.35,投影レンズ CF photo 8x)を使用し,HFM型撮影装置により撮影した.フィルムはKodak High Contrast Copy film (5069)を用い,DK-20処方で現像した.このフィルムをフィルムスキャナーにより解像度2,400 dpi で読み取りJPEG画像データとして保存した.
図版のスケールは原則として10μmであり,それ以外のものはそれぞれ示した.
花粉の計測
花粉の粒径は50個体の測定値を求め,最大値,最小値を示した.極長(polar lenght)と赤道径(equatorial diameter)の異なる場合は赤道径×極長として示した.また,極の不明瞭な花粉では最大径を,必要に応じて最小径も示した.これらの測定には接眼ミクロメーターや㈱コーガク製のデジタル測微計を使用した.
その他,突起や網目の単位面積当たりの密度も必要に応じて示した.これらは原則として単溝型(monocolpate)の花粉では向心極面中央部,その他の花粉ではmesocolpiumの5×5または10×10μm当たりの突起数を示してある.花粉外壁(exine)の厚さが一様では無いときは原則として向心極面またはmesocolpiumで測定した.
記載した花粉について
記載した種類は花粉分析の対象になりうる種類のみに限定し,ラン科(Orchidaceae),ガガイモ属(Metoplexis)など化石花粉となる可能性のきわめて低い種類は除外した.
なお,写真および花粉の記載種の学名および配列は,中村(1980)では大井(1978)に準拠していたが,今回の配列は邑田・米倉(2013)によるAPGIII分類体系の形式に,学名は米倉・梶田 (2003-)にしたがった.
花粉外壁(exine)の構造
花粉の細胞は主にペクチン(Pectin)とセルロース(cellulose)からなる内壁(intine)とスポロポレニン(sporopollenin)からなる外壁(exine)で覆われているが,acetolysis処理をすると花粉は化学的に強靱な外壁部分だけとなる.この花粉外壁(exine)は内層(endexine)と外層(ektexine*)に分かれ,
さらに外層は底部層(foot layer),柱状層(collumella),外表層(tectum)とに分けられる(図1).このような花粉外壁の層状構造は,裸子植物では認められないが,多くの被子植物ではごく普通に認められる.花粉外壁や花粉管の出口である発芽装置(発芽口)は送粉様式や進化などに関連して様々に変化した結果,花粉の形を決めている.
注):* ectexine ともいう(Punt et al., 2007)
図1:花粉外壁の層状構造
花粉の形態は,植物の種類によってそれぞれ特徴をもつが,花粉を識別する際に重要な特徴とは,1)花粉の大きさ,2)花粉の外形,3)発芽口の数,位置とその配列,形,構造,4)花粉表面にある模様(彫紋)とその構造である.
これらの形や位置も,観察する方向が決まっていなければ正確な記載ができないので,花粉には地球と同じように方向が定められている.花粉母細胞が分裂し花粉4分子が形成され,これが発育して花粉粒となる.その際,各分子は立体的あるいは平面上に配列する.この花粉粒がバラバラに離れずにいる時,その中心に向かう軸(極軸)にそって中心方向を向心極,反対方向を遠心極と呼ぶ.極軸中央部を垂直交叉する軸が赤道軸である.
1)花粉の大きさ
外形が同じであっても大きさが著しく異なる場合のように,花粉の大きさは種類を区別するための重要な性質である.形態記載には必ず粒径を示しておく必要がある.場合によっては,発芽口の大きさや花粉表面の突起などの大きさなども重要である.なお,花粉の大きさは,処理法,封入剤,封入後の時間によって変化する.一般的には,KOH法よりもacetolysis法で処理したものの方が,脱水の必要な封入剤よりも脱水の不必要な封入剤の方が,花粉は大きくなる傾向がある.処理法や封入剤の違いによる花粉粒径などの違いに関しては,Reitsma(1969),Meltsov et al.(2008)を参照のこと.今回はsilicon oilを封入剤として用いたため時間による粒径の変化は無視してよい.
2)花粉の外形
花粉粒が形成される際に個々の花粉粒が分離せず結合したままのものがある.このような花粉を多集粒花粉というが,この集合状態も識別には重要である.
裸子植物の一部には花粉に袋状の気囊とよばれる構造物をもつものがあり,その大きさ,形状,花粉本体への付着の仕方などに植物ごとの特徴がみられる.
多くの花粉は単粒であるが,その外形は見る方向により様々な形が観察される.極方向からみた場合には極観といい,その輪郭の形により様々な形が認められる.赤道方向からの場合は赤道観とよび,極方向(P)と赤道方向(E)の長さの比(P/E ratio)から過長球形〜過偏球形まで様々な形が区別される(図2).極観や赤道観は封入方法や経年によって変化することや常に理想的な方向から観察できるとは限らないことから,実際の観察時にはかなり経験に基づいた修正が必要となることも多い.
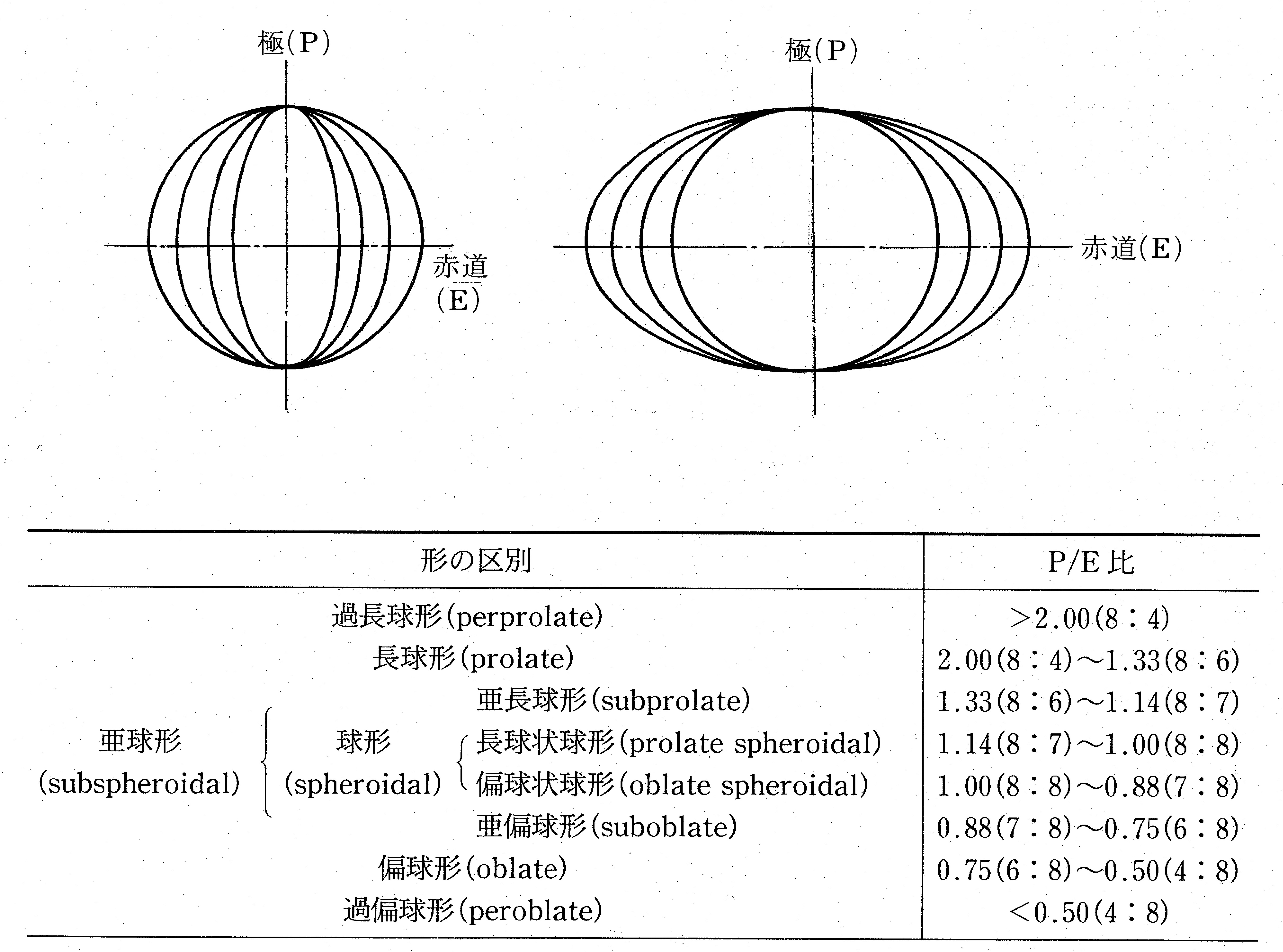
図2:Erdtman (1952) のP/E比に基づく花粉類型
3)発芽口
発芽口はその形から孔と溝に分けられる.前者は円形あるいは短径と長径の比が2以下の楕円形のものをさし,後者は短径と長径の比が2以上で多くはその両端が尖っている裂け目状のものをいう.発芽口は花粉外壁(exine)の外層(extexine)にある外口(ectoaperture)と,内層(endexine)にある内口(endoaperture)に分けられ,外口と内口の形と組み合わせから外見的には孔型,溝型,溝孔型に区別される.また,口周辺の外層と内層の構造からさらに細分されるのが普通である.発芽口の数,位置とその配列,形,構造は分類群ごとにほぼ決まっているので重要な特徴である.発芽口が複数ある花粉の多くは赤道面上に配列するか赤道面に対称に配列するが,なかにはカヤツリグサ科のように異極性(heteropolar)を示すものもある.なお,裸子植物では遠心極面の花粉外壁は薄いことが多く,この部分が,発芽に際して花粉管の出る場所となる(薄膜部:leptoma).
4)花粉表面にある彫紋とその構造
彫紋には様々な型が見られるが,これらは花粉外壁の外層断面の構造や構成要素(エレメント)に基づいている.外層のうち柱状層(columella)より上部に外表層(tectum)をもつ花粉(外表層型:tectate)ともたない花粉(外表層欠如型:intectate)があり,前者の場合には最上部に合焦したときと柱状層に合焦したときでは異なる彫紋が観察されることも多い.したがって,彫紋の観察には,花粉壁断面をよく観察するとともに焦点移動によるL-O分析が重要となる(Erdtman, 1952).
検索表および記載用語について
花粉は様々な形や構造をもつため,それらを記載する用語が多くの花粉学者によって定義されている.ここでの花粉型の分類は,Faegri and Iversen(1975)にしたがい24類型を用いた.また,形態記載用語は「花粉学事典」およびPunt et al.,(2007)の "Glossary of pollen and spore terminology"に準拠した.
終わりに,出版にあたって様々な援助をいただいた岡山理科大学 三好教夫名誉教授および韓国蔚山大学 崔 基龍(Kee-Ryong Choi)教授ならびに大阪自然史博物館の学芸員諸氏に厚く御礼を申し上げる.
引用文献
Andersen, S. Th.(1960)Silicone Oil as a Mounting Medium for Pollen Grains. Danmarks Geologiske Undersøgelse IV. Række, vol. 4, 1–24.
Erdtman, G(1943)An introduction to Pollen Analysis. Chronica botanica Company, Waltham.
Erdtman, G. (1952) Pollen Morphology and Plant Taxonomy. Almqvist & Wiksell, Stockholm.
Faegri, K.and J. Iversen (1975) Textbook of modern pollen analysis. 3rd Ed. Munksgaard, Copenhagen.
Jimbo, T.(1933): The Diagnoses of the pollen of Forest Trees. 1. Sci. Rep.Tohoku Univ 4th. ser. (Biol), vol.8 287-299.
Larson, D. A., Skvarla, J. J., and Lewis, C. W., (1962): An electron microscope study of exine stratification and fine structure. Pollen et Spores, vol. 4, 233–246.
Meltsov, V., Poska, A. & Saar, M.(2008): Pollen size in Carex: The effect of different chemical treatments and mounting media. Grana, vol. 47, 220-233.
中村 純(1980)日本産花粉の標徴 I・II.大阪市立自然史博物館収蔵資料目録第12・13集, 大阪市立自然史博物館,大阪.
日本花粉学会(1994)花粉学事典.朝倉書店,東京.
大井 次三郎(1978)日本植物誌 顕花篇 改訂増補版.至文堂,東京.Revised and enlarged edition.
Punt W, Hoen PP, Blackmore S, Nilsson S, Le Thomas A. (2007) Glossary of pollen and spore terminology. Review of Palaeobotany and Palynology, vol. 143, 1–81.
Reitsma, T. (1969) Size modification of recent pollen grains under different treatments. Review of Palaeobotany and Palynology, 9, 175–202.
山崎 次男(1933)樺太演習林所産植物 花粉並ニ胞子形態.京都帝國大學演習林報告 第五號.
米倉 浩司・梶田 忠(2003-)BG Plants 和名-学名インデックス(YList),http://ylist.info/
米倉 浩司, 邑田 仁(2013)維管束植物分類表.北隆館,東京.
--------------------------------------------------